
rongsheng jin lab quotation
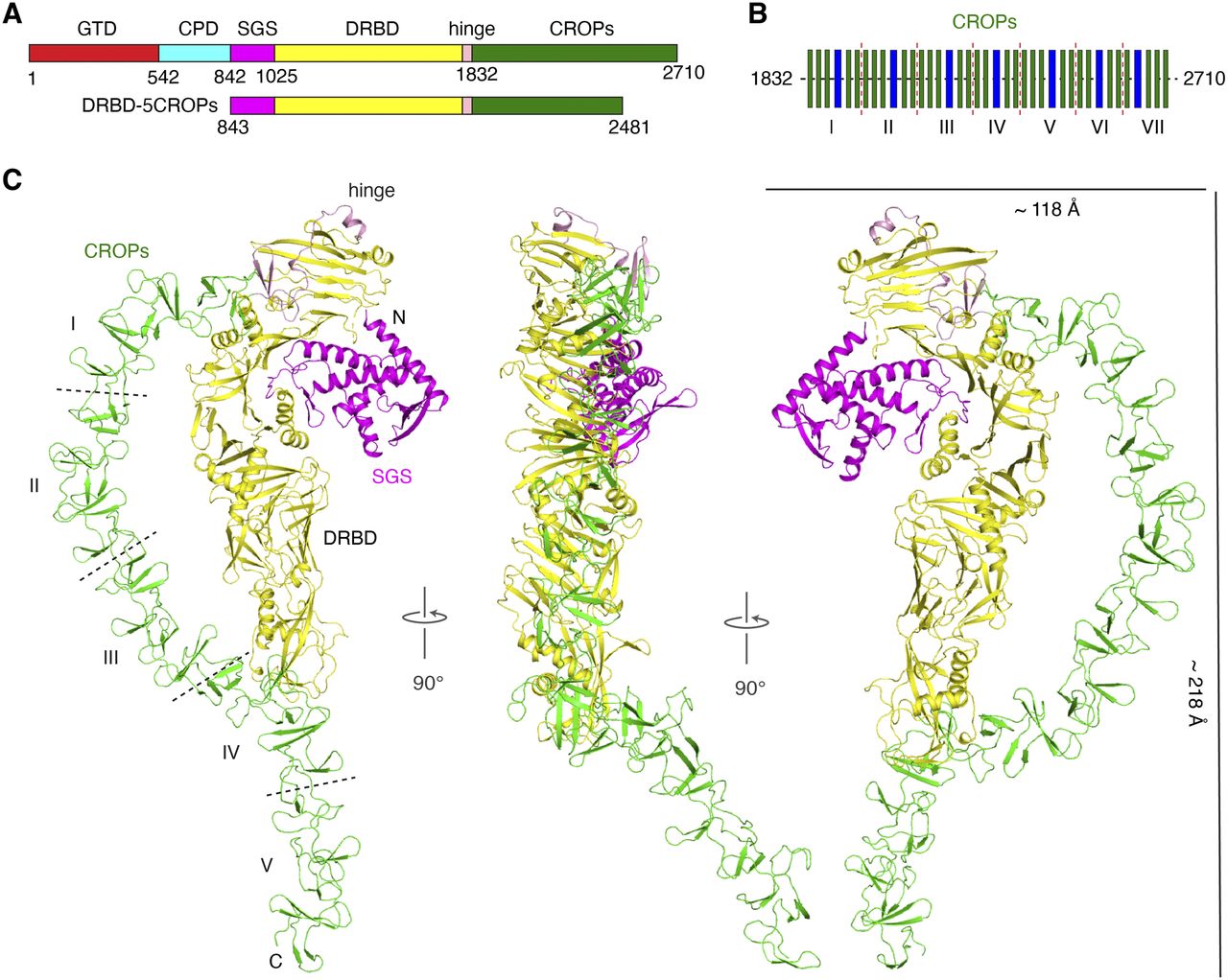
Lam, K. H., Guo, Z., Krez, N., Matsui, T., Perry, K., Weisemann, J., Rummel, A., Bowen, M. E. & Jin, R. A viral-fusion-peptide-like molecular switch drives membrane insertion of botulinum neurotoxin A1. Nat Commun 9, 5367 (2018) doi: 10.1038/s41467-018-07789-4.
Chen, P., Tao, L., Liu, Z., Dong, M. & Jin, R. Structural insight into Wnt signaling inhibition by Clostridium difficile toxin B. FEBS J (2018) doi: 10.1111/febs.14681.
Chen, P., Tao, L., Wang, T., Zhang, J., He, A., Lam, K. H., Liu, Z., He, X., Perry, K., Dong, M*. & Jin, R*. Structural basis for recognition of frizzled proteins by Clostridium difficile toxin B. Science 360, 664-669 (2018) (*corresponding authors) doi: 10.1126/science.aar1999. PMCID: PMC6231499
Lam, K. H., Sikorra, S., Weisemann, J., Maatsch, H., Perry, K., Rummel, A., Binz, T. & Jin, R. Structural and biochemical characterization of the protease domain of the mosaic botulinum neurotoxin type HA. Pathog Dis 76 (2018) doi: 10.1093/femspd/fty044. PMCID: PMC5961070
Silva, D. A., Stewart, L., Lam, K. H., Jin, R. & Baker, D. Structures and disulfide cross-linking of de novo designed therapeutic mini-proteins. FEBS J 285, 1783-1785 (2018) doi: 10.1111/febs.14394. PMCID: PMC6001749
Lam, K. H., Qi, R., Liu, S., Kroh, A., Yao, G., Perry, K., Rummel, A. & Jin, R. The hypothetical protein P47 of Clostridium botulinum E1 strain Beluga has a structural topology similar to bactericidal/permeability-increasing protein. Toxicon 147, 19-26 (2018) doi: 10.1016/j.toxicon.2017.10.012. PMCID: PMC5902665
Chevalier, A., Silva, D.A., Rocklin, G.J., Hicks, D.R., Vergara, R., Murapa, P., Bernard, S.M., Zhang, L., Lam, K.H., Yao, G., Bahl, C.D., Miyashita, S.I., Goreshnik, I., Fuller, J.T., Koday, M.T., Jenkins, C.M., Colvin, T., Carter, L., Bohn, A., Bryan, C.M., Fernández-Velasco, D.A., Stewart, L., Dong, M., Huang, X., Jin, R., Wilson, I.A., Fuller, D.H. & Baker, D. Massively parallel de novo protein design for targeted therapeutics. Nature 550(7674):74-79 (2017) doi: 10.1038/nature23912. PMCID: PMC5802399
Yao, G., Lam, K.H., Weisemann, J., Peng, L., Krez, N., Perry, K., Shoemaker, C.B., Dong, M., Rummel, A. & Jin, R. A camelid single-domain antibody neutralizes botulinum neurotoxin A by blocking host receptor binding. Sci Rep. 7;7(1):7438. (2017) doi: 10.1038/s41598-017-07457-5. PMCID: PMC5547058
Yao, G., Lam, K.H., Perry, K., Weisemann, J., Rummel, A. & Jin, R. Crystal Structure of the Receptor-Binding Domain of Botulinum Neurotoxin Type HA, Also Known as Type FA or H. Toxins (Basel) 9, 93 (2017) doi: 10.3390/toxins9030093. PMCID: PMC5371848
Yao, G., Zhang, S., Mahrhold, S., Lam, K. H., Stern, D., Bagramyan, K., Perry, K., Kalkum, M., Rummel, A.*, Dong, M.* & Jin, R.* N-linked glycosylation of SV2 is required for binding and uptake of botulinum neurotoxin A. Nat Struct Mol Biol 23 (7):656-662 (2016) (*corresponding authors) doi: 10.1038/nsmb.3245. PMCID: PMC5033645
Lee, K., Lam, K. H., Kruel, A. M., Mahrhold, S., Perry, K., Cheng, L. W., Rummel, A. & Jin, R. Inhibiting oral intoxication of botulinum neurotoxin A complex by carbohydrate receptor mimics. Toxicon 107, 43-49 (2015) doi: 10.1016/j.toxicon.2015.08.003. PMCID: PMC4658216
Lam, K.H. & Jin, R. Architecture of the botulinum neurotoxin complex: a molecular machine for protection and delivery. Current Opinion in Structural Biology 31:89-95 (2015) doi: 10.1016/j.sbi.2015.03.013. PMCID: PMC4476938
Lam, K.H., Yao, G. & Jin, R. Diverse binding modes, same goal: The receptor recognition mechanism of botulinum neurotoxin. Progress in Biophysics and Molecular Biology 117(2-3):225-31 (2015) doi: 10.1016/j.pbiomolbio.2015.02.004. PMCID: PMC4417461
Lam, T.I., Stanker, L.H., Lee, K., Jin, R. & Cheng, L.W. Translocation of botulinum neurotoxin serotype A and associated proteins across the intestinal epithelia. Cellular Microbiology 17(8):1133-1143 (2015) doi: 10.1111/cmi.12424. PMCID: PMC4610714
Matsui, T.*, Gu, S., Lam, K.H., Carter, L.G., Rummel, A., Mathews, II. & Jin, R.* Structural Basis of the pH-Dependent Assembly of a Botulinum Neurotoxin Complex. J. Mol. Biol. 426(22):3773-3782 (2014) doi: 10.1016/j.jmb.2014.09.009. (*corresponding authors) PMCID: PMC4252799
Lee, K., Zhong, X., Gu, S., Kruel, A.M., Dorner, M.B., Perry, K., Rummel, A., Dong, M. & Jin, R. Molecular basis for disruption of E-cadherin adhesion by botulinum neurotoxin A complex. Science 344(6190):1405-1410 (2014) doi: 10.1126/science.1253823. PMCID: PMC4164303
Lee, K., Lam, K.H., Kruel, A.M., Perry, K., Rummel, A. and Jin, R. High-resolution crystal structure of HA33 of botulinum neurotoxin type B progenitor toxin complex. Biochem. Biophys. Res. Commun. 446(2):568-573 (2014) doi: 10.1016/j.bbrc.2014.03.008. PMCID: PMC4020412
Yao, Y., Lee, K., Gu, S., Lam, K.H. & Jin, R. Botulinum Neurotoxin A Complex Recognizes Host Carbohydrates through Its Hemagglutinin Component, Toxins (Basel) 6(2):624-635 (2014) doi: 10.3390/toxins6020624. PMCID: PMC3942755
Lee, K., Gu, S., Jin, L., Le, T.T.N., Cheng, L.W., Strotmeier, J., Kruel, A.M., Yao, G., Perry, K., Rummel, A.* & Jin, R.* Structure of a Bimodular Botulinum Neurotoxin Complex Provides Insights into Its Oral Toxicity. PLoS Pathog. 9(10): e1003690 (2013) doi:10.1371/journal.ppat.1003690. (*corresponding authors) PMCID: PMC3795040
Zong, Y. and Jin, R. Structural mechanisms of the agrin-LRP4-MuSK signaling pathway in neuromuscular junction differentiation. Cell. Mol. Life Sci. 70(17):3077-88 (2013) doi: 10.1007/s00018-012-1209-9. PMCID: PMC4627850
Gu, S. and Jin, R. Assembly and function of the botulinum neurotoxin progenitor complex. Curr. Top. Microbiol. Immunol. 364:21-44 (2013) doi: 10.1007/978-3-642-33570-9_2. PMCID: PMC3875173
Gu, S., Rumpel, S., Zhou, J., Strotmeier, J., Bigalke, H., Perry, K., Shoemaker, C.B., Rummel, A. & Jin, R. Botulinum neurotoxin is shielded by NTNHA in an interlocked complex. Science 335(6071):977-81 (2012) doi: 10.1126/science.1214270. PMCID: PMC3545708
Zong, Y., Zhang, B., Gu, S., Lee, K., Zhou, J., Yao, G., Figueiredo, D., Perry, K., Mei, L.* & Jin, R.* Structural basis of neuron-specific regulation of postsynaptic differentiation. Gene & Development 26:247-258 (2012) doi: 10.1101/gad.180885.111. (*corresponding authors) PMCID: PMC3278892
Yao, G., Zong, Y., Gu, S., Zhou, J., Xu, H., Mathews, II. & Jin, R. Crystal structure of the glutamate receptor GluA1 amino-terminal domain. Biochem. J. 438(2):255-63 (2011) doi: 10.1042/BJ20110801. PMCID: PMC3296483
Strotmeier, J., Gu, S., Jutzi, S., Mahrhold, S., Zhou, J., Pich, A., Eichner, T., Bigalke, H., Rummel, A.*, Jin, R.* & Binz, T*. The biological activity of botulinum neurotoxin type C is dependent upon novel types of ganglioside binding sites. Mol. Microbiol. 81(1):143-56 (2011) doi: 10.1111/j.1365-2958.2011.07682.x. Epub 2011 Jun 2. (*corresponding authors)
Strotmeier, J., Lee, K., Völker, A.K., Mahrhold, S., Zong, Y., Zeiser, J., Zhou, J., Pich, A., Bigalke, H., Binz, T., Rummel, A.* & Jin, R.* Botulinum neurotoxin serotype D attacks neurons via two carbohydrate-binding sites in a ganglioside-dependent manner. Biochem. J. 431(2):207-16 (2010) (*corresponding authors)
Jin, R.*, Singh, S.K., Gu, S., Furukawa, H., Sobolevsky, A.I., Zhou, J., Jin, Y. & Gouaux E.* Crystal structure and association behavior of the GluR2 amino-terminal domain. EMBO J. 28(12):1812-23 (2009) (*corresponding authors) PMCID: PMC2699365
Kumar, J., Schuck. P., Jin, R. & Mayer, M.L. The N-terminal domain of GluR6-subtype glutamate receptor ion channels. Nat. Struct. Mol. Biol. 16(6):631-8 (2009) PMCID: PMC2729365
Jin, R., Rummel, A., Binz, T. & Brunger, A.T. Botulinum neurotoxin B recognizes its protein receptor with high affinity and specificity. Nature 444:1092-5 (2006)
Jin, R., Clark, S., Weeks, A.M., Dudman, J.T., Gouaux, E. & Partin, K.M. Mechanism of positive allosteric modulators acting on AMPA receptors. J. Neurosci. 25(39):9027-36 (2005)
Jin, R., Junutula, J.R., Matern, H.T., Ervin, K.E., Scheller, R.H. & Brunger, A.T. Exo84 and Sec5 are competitive regulatory Sec6/8 effectors to the RalA GTPase. EMBO J. 24:2064-74 (2005)
Jin, R., Bank, T., Mayer, M. L., Traynelis, S. & Gouaux, E. Structural basis for partial agonist action at ionotropic glutamate receptors. Nat. Neurosci. 6(8):803-10 (2003)

Jahid S, Ortega JA, Vuong LM, Acquistapace IM, Hachey SJ, Flesher JL, La Serra MA, Brindani N, La Sala G, Manigrasso J, Arencibia JM, Bertozzi SM, Summa M, Bertorelli R, Armirotti A, Jin R, Liu Z, Chen CF, Edwards R, Hughes CCW, De Vivo M, Ganesan AK. PMID: 35385746; PMCID: PMC9127750.
Chen P, Zeng J, Liu Z, Thaker H, Wang S, Tian S, Zhang J, Tao L, Gutierrez CB, Xing L, Gerhard R, Huang L, Dong M, Jin R. PMID: 34145250; PMCID: PMC8213806.
Chen P, Lam KH, Liu Z, Mindlin FA, Chen B, Gutierrez CB, Huang L, Zhang Y, Hamza T, Feng H, Matsui T, Bowen ME, Perry K, Jin R. PMID: 31308519; PMCID: PMC6684407.

Published in Cell Reports, the paper is titled, “Structural insights into rational design of single-domain antibody-based antitoxins against botulinum neurotoxins.” Led by Rongsheng Jin, PhD, a professor in the Department of Physiology & Biophysics at the University of California, Irvine, School of Medicine, this paper describes how the team first identified the neutralizing epitopes of six anti-BoNT nanobodies (VHHs) based on their crystal structures, then harnessed the structural findings to rationally design bifunctional nanobodies. Different than ordinary nanobodies, bifunctional nanobodies are composed of two nanobodies that bind simultaneously to the toxins.
“In a nutshell, we establish a platform for structure-based rational design of bifunctional antitoxins against BoNTs,” said Kwok-ho Lam, the first author and a project scientist in the Jin lab. “BoNTs can be misused as a bioweapon and thus have been classified as Tier 1 select agents by the Centers for Disease Control and Prevention, which is why there is urgent need for antitoxins.”
“Currently, the only available antitoxin remedies are polyclonal antibodies from horse or human serum, which have known health risks and are in limited supply. Monoclonal antibodies are still under development,” said Jin. “And, while it isn’t necessarily a cause for worry, the increasingly popular therapeutic uses of BoNT products also create risks of possible botulism resulting from the medical treatments where they are used.”
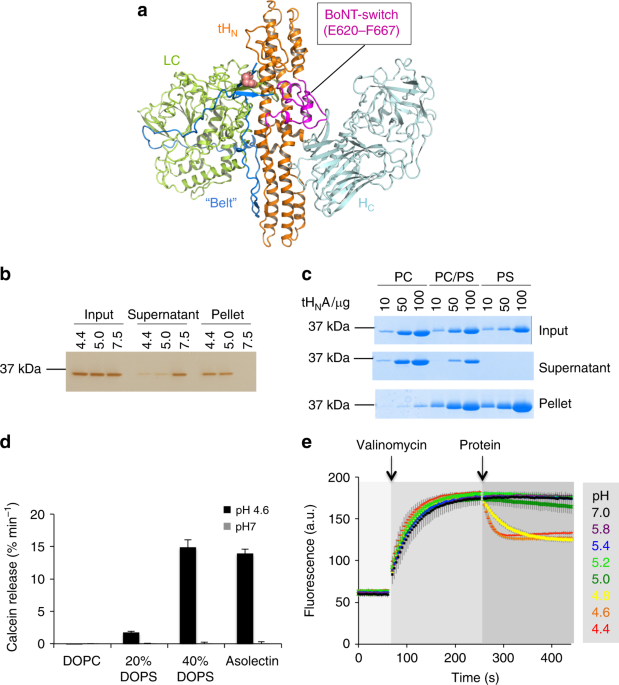
Wild type and mutated recombinant full-length activated BoNT/A1 were produced under biosafety level 2 containment (project number GAA A/Z 40654/3/123) recombinantly in K12 E. coli strain in Dr. Rummel’s lab6-tag were purified on Co2+-Talon matrix (Takara Bio Europe S.A.S., France) and eluted with 50 mM Tris-HCl (pH 8.0), 150 mM NaCl, and 250 mM imidazole. For proteolytic activation, BoNT/A1 was incubated for 16 h at room temperature with 0.01 U bovine thrombin (Sigma-Aldrich Chemie GmbH, Germany) per µg of BoNT/A1. Subsequent gel filtration (Superdex-200 SEC; GE Healthcare, Germany) was performed in phosphate buffered saline (pH 7.4).
The purified ciA-D12 (S124C) was labeled with Alexa Fluor C2 647 maleimide (Molecular Probes) according to the manufacturer’s instructions. The labeled ciA-D12 was further purified by MonoQ ion-exchange chromatography in 10 mM Hepes (pH 8.0) and eluted with a NaCl gradient. The calculated dye to protein ratio was ~1 mole of dye per mole of ciA-D12. BoNT/A1i-ciA-D12 complex were prepared by mixing BoNT/A1i and ciA-D12 in 1:1.5 molar ratio and the complex was purified by size-exclusion chromatography using Superdex-200.
Liposomes containing 10% GT1b, 69% DOPC, 20% DOPS, 0.5% rhodamine-PE, and 0.5% biotin-PE were prepared by extrusion through a 100 nm pore membrane. To form proteoliposomes, 10 nM BoNT/A1i–D12* or oxidized BoNT/A1iDS–D12* was incubated with 0.5 mg/ml lipid at room temperature for 1 h at the pH indicated. The mixture was then diluted 1000-fold and incubated for 5 minutes in a passivated, quartz microscope chamber functionalized with streptavidin. The biotinylated liposomes were retained and unbound proteins are washed away by exhaustive rinsing with buffer. At the low densities needed for optical resolution of individual liposomes, we could observe sufficient liposomes for statistical analysis, while minimizing the probability that a diffraction-limited spot would contain multiple liposomes. Samples were imaged using a prism-based Total Internal Reflection Fluorescence (TIRF) microscope. Samples were first excited with a laser diode at 640 nm (Newport Corporation, Irvine, CA) to photobleach Alexa 647-labeled BoNT/A1i–D12* or BoNT/A1iDS-D12* molecules followed by excitation with a diode pumped solid state laser at 532 nm (Newport Corporation, Irvine, CA) to probe for Rhodamine-labeled liposomes. Emission from protein and lipids was separated using an Optosplit ratiometric image splitter (Cairn Research Ltd., Faversham UK) containing a 645 nm dichroic mirror, a 585/70 band pass filter for Rhodamine, and a 670/30 band pass filter for Alexa 647 (all Chroma, Bellows Falls, VT). The replicate images were relayed to a single iXon EMCCD camera (Andor Technologies, Belfast, UK) at a frame rate of 10 Hz. Data were processed in MATLAB to cross-correlate the replicate images and extract time traces for diffraction-limited spots with intensity above baseline. Single-molecule traces were hand selected based on the exhibition of single-step decays to baseline during 640 illumination and the appearance of Rhodamine emission during 532 illumination.
Citation:Lee K, Gu S, Jin L, Le TTN, Cheng LW, Strotmeier J, et al. (2013) Structure of a Bimodular Botulinum Neurotoxin Complex Provides Insights into Its Oral Toxicity. PLoS Pathog 9(10):
Structural information of HAs is available for serotypes C and D, such as the crystal structures of HA33 of serotype C (HA33-C) [12], [13], a complex composed of HA17 and HA33 of serotype D [14], and HA70 of serotype C (HA70-C) [15], [16]. However, BoNT/C and D rarely cause human botulism but are known to cause the syndrome in cattle, poultry, and wild birds. For BoNT/A, the major cause of human botulism, only the structure of HA33 (HA33-A), which displays an amino-acid identity of ∼38% to HA33-C and D, has been solved [17]. We have recently determined the crystal structure of the BoNT/A–NTNHA complex [5]. However, it remains largely unclear how the HAs assemble with one another and how they interact with BoNT and NTNHA.
Interacting residues are labeled in open-book views of the interfaces. (A) Interface between HA70 and HA17. (B) Interfaces between HA17 and the two HA33s are indicated by purple and blue. The HA33 residues involved in both interfaces are in green. (C) Interface between two HA33s attached to the same HA17. See Fig. S5 and S6 in Text S1 for stereo versions of the detailed interactions.
To determine the physiological relevance of these HA–glycan interactions during toxin absorption, we examined their ability to interfere with the HA complex-mediated disruption of Caco-2 TER. Lac, Gal, and IPTG markedly inhibited the TER reduction induced by the HA complex, and showed higher potencies when applied to the apical than to the basolateral compartment (Fig. 5A–B and Fig. S11A–D in Text S1). In contrast, α2,3- and α2,6-SiaLac, and to a lesser extent Neu5Ac, inhibited the decrease in TER only when applied apically, albeit more weakly than Lac (Fig. 5A–B and Fig. S11E–F in Text S1). We then examined the transport of the HA complex across the Caco-2 monolayer using a fluorescence-labeled HA complex (HA*) (Fig. 5C). Lac and IPTG efficiently inhibited the transport of HA* when it was applied to the apical or basolateral chamber. Blocking the transport of HA* via α2,3- and α2,6-SiaLac was more potent toward the basolateral compartment than toward the apical side. Neu5Ac at 50 mM did not inhibit transport of HA* from either side of the Caco-2 cell monolayer. These data are consistent with the ability of these carbohydrates to inhibit TER reduction induced by the HA complex. Collectively, these results suggest that the binding of HAs to Neu5Ac- and Gal-containing glycans on epithelial cells is essential for the transport of BoNT across the intestinal wall. Moreover, the carbohydrate receptors may play a more important role in the initial L-PTC absorption in the intestinal lumen, whereas other host receptors (e.g., E-cadherin) are involved once it gains access to the basolateral side.
(A, B) TER of Caco-2 monolayers was measured when Alexa-488-labeled HA complex (HA*) pre-incubated with Lac, IPTG, α2,3-SiaLac, or α2,6-SiaLac was applied to the apical (A; 58 nM) or basolateral (B; 17 nM) chambers. Values are means ± SD (n = 4–12). (C) HA* (with or without carbohydrates) or the Alexa-488-labeled HA33-DAFA complex (HA33-DAFA *) was applied to the apical (at 58 nM) or basolateral (at 17 nM) chamber. The fluorescence signals in both chambers were quantified after 24 hours and the amount of transported HA*/HA33-DAFA * was expressed as a percentage of the total HA*/HA33-DAFA * used. Values are means ± SD (n = 3–22).
The loose linkage between the M-PTC and the HA complex clearly suggests divided functions. We previously reported that the M-PTC"s compact structure protects BoNT against digestive enzymes and the extreme acidic environment of the GI tract [5], [23]. We now show that the HA complex is mainly responsible for BoNT absorption in the small intestine, through binding to specific host carbohydrate receptors. This new finding permitted the identification of IPTG as a prototypical oral inhibitor that extends survival following lethal oral BoNT/A intoxication of mice. Multivalent interactions involving nine binding sites for Neu5Ac- and Gal-containing glycans increase the overall avidity of binding between the L-PTC and glycans on the epithelial cell surface, and thus compensate for the modest glycan-binding affinities at individual binding sites (Fig. 6C). Similarly, the potency of carbohydrate receptor mimics could be improved by optimizing the HA–glycan interactions as revealed here or by introducing new HA–inhibitor interactions at individual binding sites based on rational design, as well as by designing multivalent inhibitors. Although such inhibitors cannot be used to treat fully developed food-borne botulism, they could provide temporary protection upon pre-treatment and could also be useful for cases of intestinal colonization with C. botulinum spores such as in cases of infant or adult intestinal botulism. Our results also suggest that the L-PTC could be exploited for alternative applications. For example, protein-based therapeutics could be coupled to the modified non-toxic L-PTC to allow oral delivery by improving drug stability, absorption efficiency, and bioavailability.
The purified HA70 was labeled with Alexa Fluor® 488 carboxylic acid, succinimidyl ester (Life Technologies) according to the manufacturer"s instructions. The labeled HA70 was further purified by Superdex 200 chromatography in 20 mM Tris (pH 8.0) and 50 mM NaCl. The calculated dye to protein ratio was ∼2 moles of dye per mole of monomeric HA70.
The HA17–HA33, the HA70–HA17, and the HA70D3–HA17 complexes were produced by co-expression and co-purification as described above. To assemble the mini-HA complex (HA70D3–HA17–HA33), the purified HA33 and the HA70D3–HA17 complex were mixed at a molar ratio of ∼2.5∶1 and incubated at 4°C overnight. The excess HA33 was removed by Superdex 200 chromatography with 20 mM Tris (pH 7.6) and 50 mM NaCl. The fully assembled HA complex was reconstituted by mixing the purified HA70 and the HA17–HA33 complex at a molar ratio of ∼1∶1.3. The mixture was incubated at 4°C overnight and the excess HA17–HA33 complex was removed from the mature HA complex by Superdex 200 chromatography with 20 mM Tris (pH 7.6) and 50 mM NaCl. The fluorescence-labeled HA complex was prepared with Alexa Fluor® 488-labeled HA70 and unlabeled HA17–HA33 complex (HA*) or HA17–HA33DAFA complex (HA33DAFA*) using a similar protocol.
Sedimentation equilibrium (SE) experiments were performed in a ProteomeLab XL-I (BeckmanCoulter) analytical ultracentrifuge. Purified HA samples were dialyzed extensively against a buffer containing 50 mM Tris (pH 7.6) and various NaCl concentrations, or 50 mM citric acid (pH 2.3) and various NaCl concentrations. Protein samples at concentrations of 0.4, 0.2, and 0.1 unit of OD280 were loaded in 6-channel equilibrium cells and centrifuged at 20°C in an An-50 Ti 8-place rotor at the first speed indicated until equilibrium was achieved and thereafter at the second speed. HA33 was analyzed at rotor speeds of 19,000 and 22,000 rpm. The HA17–HA33 and the HA70D3–HA17–HA33 complexes were analyzed at 12,000 and 14,000 rpm. The HA70–HA17 and the HA70–HA17–HA33 complexes were run at speeds of 6,000 and 8,000 rpm. For each sample, data sets for the two different speeds were analyzed independently using HeteroAnalysis software (by J.L. Cole and J.W. Lary, University of Connecticut). Three independent experiments were performed for each sample.
The L-PTC of BoNT/A was obtained from List Biological Laboratories, Inc. (Campbell, California) and Miprolab GmbH (Göttingen, Germany). The recombinant HA complex was reconstituted in vitro as described above. Negatively stained EM specimens were prepared following a previously described protocol [54]. Briefly, 3 µl of the L-PTC (∼0.02 mg/ml in 20 mM MES, pH 6.2, and 100 mM NaCl) or the HA complex (∼0.01 mg/ml in 20 mM Tris, pH 7.6, and 50 mM NaCl) was placed on a freshly glow-discharged carbon-coated EM grid, blotted with filter paper after 40 seconds, washed with two drops of deionized water, and then stained with two drops of freshly prepared 1% uranyl formate, which also served to fix the proteins.
Carbohydrate inhibition assays: Lac, Gal, IPTG, Neu5Ac, α2,6- and α2,3-SiaLac were dissolved in IMDM, sterile filtered and stored at −20°C. Neu5Ac stock solution was adjusted to pH 7.4. The wild type HA complex (HA wt), fluorescence-labeled HA complex (HA*), or the HA70-TPRA complex were pre-incubated with the corresponding carbohydrate over night at 4°C in IMDM and diluted to the final concentration with IMDM prior to administration. The TER upon administration of each carbohydrate in the highest concentrations used was checked in the absence of HA and was virtually identical to that of the control without sugars.
Transport measurement: For paracellular transport studies, filters were incubated in IMDM added to the apical (0.5 ml) and basolateral (1.5 ml) reservoir. As marker substance Alexa Fluor® 488 labeled HA* or HA33-DAFA* was administered to the apical or basolateral reservoirs at final concentrations of 58 nM and 17 nM, respectively. After 24 hour of incubation, 200 µl of samples were taken from the apical and the basolateral reservoir. The marker substance was measured in a BioTek Synergy 4 fluorescence spectrophotometer at 495 nm excitation and 519 nm emission wavelengths.
4.Cheng LW, Onisko B, Johnson EA, Reader JR, Griffey SM, et al. (2008) Effects of purification on the bioavailability of botulinum neurotoxin type A. Toxicology 249: 123–129.
20.Inoue K, Fujinaga Y, Watanabe T, Ohyama T, Takeshi K, et al. (1996) Molecular composition of Clostridium botulinum type A progenitor toxins. Infect Immun 64: 1589–1594.
25.Fujinaga Y, Inoue K, Nomura T, Sasaki J, Marvaud JC, et al. (2000) Identification and characterization of functional subunits of Clostridium botulinum type A progenitor toxin involved in binding to intestinal microvilli and erythrocytes. FEBS Lett 467: 179–183.
26.Inoue K, Fujinaga Y, Honke K, Arimitsu H, Mahmut N, et al. (2001) Clostridium botulinum type A haemagglutinin-positive progenitor toxin (HA(+)-PTX) binds to oligosaccharides containing Gal beta1-4GlcNAc through one subcomponent of haemagglutinin (HA1). Microbiology 147: 811–819.
27.Sugawara Y, Matsumura T, Takegahara Y, Jin Y, Tsukasaki Y, et al. (2010) Botulinum hemagglutinin disrupts the intercellular epithelial barrier by directly binding E-cadherin. J Cell Biol 189: 691–700.
28.Jin Y, Takegahara Y, Sugawara Y, Matsumura T, Fujinaga Y (2009) Disruption of the epithelial barrier by botulinum haemagglutinin (HA) proteins - differences in cell tropism and the mechanism of action between HA proteins of types A or B, and HA proteins of type C. Microbiology 155: 35–45.
29.Matsumura T, Jin Y, Kabumoto Y, Takegahara Y, Oguma K, et al. (2008) The HA proteins of botulinum toxin disrupt intestinal epithelial intercellular junctions to increase toxin absorption. Cell Microbiol 10: 355–364.
32.Fujita R, Fujinaga Y, Inoue K, Nakajima H, Kumon H, et al. (1995) Molecular characterization of two forms of nontoxic-nonhemagglutinin components of Clostridium botulinum type A progenitor toxins. FEBS Lett 376: 41–44.
33.Ohyama T, Watanabe T, Fujinaga Y, Inoue K, Sunagawa H, et al. (1995) Characterization of nontoxic-nonhemagglutinin component of the two types of progenitor toxin (M and L) produced by Clostridium botulinum type D CB-16. Microbiol Immunol 39: 457–465.
Botulinum neurotoxins (BoNTs) are extremely toxic to humans by causing flaccid paralysis of botulism. The catalytic light chain (LC) of BoNTs is the warhead of the toxin, which is mainly responsible for BoNT’s neurotoxic effects. As an endopeptidase, LC is delivered by the toxin to inside neurons where it specifically cleaves neuronal SNARE proteins and causes muscle paralysis. While the currently available equine and human antitoxin sera can prevent further intoxication, they do not promote recovery from paralysis that has already occurred. We strike to develop single-domain variable heavy-chain (VHH) antibodies targeting the LC of BoNT/A (LC/A) and BoNT/B (LC/B) as antidotes to inhibit or eliminate the intraneuronal LC protease. Here, we report the identification and characterization of large panels of new and unique VHHs that bind to LC/A or LC/B. Using a combination of X-ray crystallography and biochemical assays, we reveal that VHHs exploit diverse mechanisms to interact with LC/A and LC/B and inhibit their protease activity, and such knowledge can be harnessed to predict their specificity towards different toxin subtypes within each serotype. We anticipate that the new VHHs and their characterization reported here will contribute to the development of improved botulism therapeutics having high potencies and broad specificities.
Citation:Lam K-h, Tremblay JM, Perry K, Ichtchenko K, Shoemaker CB, Jin R (2022) Probing the structure and function of the protease domain of botulinum neurotoxins using single-domain antibodies. PLoS Pathog 18(1):
Data Availability:Atomic coordinates and structure factors for sLC/A–JPU-A5–ALc-H7–JPU-C1–JPU-D12–ciA-F12, fLC/A–ALc-B8–JPU-C10–JPU-G3–JPU-D12–ciA-F12–ciA-D12, fLC/A–JPU-B9–JPU-A11–JPU-G11, sLC/B–JLJ-G3–JNE-B10, fLC/B–JSG-C1 have been deposited in the Protein Data Bank under accession codes 7L6V, 7M1H, 7LZP, 7T5F, and 7NA9, respectively. All other relevant data are within the manuscript and its Supporting Information files.
Funding:This work was partly supported by National Institute of Allergy and Infectious Diseases (NIAID) grants R01AI139087, R21AI139690, and R21AI123920 to R.J.; R01AI125704 to R.J. and C.B.S. NE-CAT at the Advanced Photon Source (APS) is supported by a grant from the National Institute of General Medical Sciences (P41 GM103403). Use of the APS, an Office of Science User Facility operated for the U.S. Department of Energy (DOE) Office of Science by Argonne National Laboratory, was supported by the U.S. DOE under Contract No. DE-AC02-06CH11357. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Botulinum neurotoxins (BoNTs) are extremely potent toxins to humans and cause flaccid paralysis. These toxins are widely distributed in nature and relatively simple to produce in quantity. For these reasons, BoNTs are listed by the Centers of Disease Control and Prevention (CDC) as Tier 1 Select Agents. BoNTs also are highly variable in nature with at least seven major serotypes reported (A-G), most of which forms a group with multiple variant subtypes. This extreme natural variability seriously complicates the development of botulism treatments. Currently the only available therapeutics for botulism are equine or human antitoxin serum products which are administered intravenously to patients exposed to toxin. Efforts are well underway to develop monoclonal antibody (mAb) antitoxins with the goal to replace current antiserum products [1–5]. While antitoxin treatments prevent further intoxication, they do not promote recovery from paralysis that has already occurred.
The full length LC/A, fLC/A, was previously reported to be unsuitable for protein crystallization because of a tendency to aggregate and the natural flexibility of its C-terminal peptide (residues 421–438) [17]. This is why all currently available crystal structures of LC/A are based on the truncated LC/A. However, the C-terminus of fLC/A is considered functionally important as it has been reported to play roles in subcellular localization [18], persistence in neurons [19,20], and catalytic activity [21]. Therefore, it was exciting to identify JPU-C10 and JPU-B9 as VHHs that bind to fLC/A, but not to sLC/A, indicating the direct involvement of the C-terminus of LC/A in VHH binding and possibly in protease activity.
In order to better understand the function of this important C-terminal region on fLC/A, we determined the structures of fLC/A in complex with these two VHHs. Surprisingly, the C-terminal region of fLC/A adopts two different conformations in these two VHH-bound complexes, further supporting the hypothesis that this region possesses natural flexibility in solution (Fig 1B and 1C). The two different conformations of fLC/A appear to have been captured as a result of their binding to VHHs. This conformational capture is not surprising as VHHs are widely used for mechanistic studies of the dynamic conformational states of enzymes and membrane proteins [22]. In the JPU-B9-bound fLC/A, the C-terminal peptide forms a loop and a short β-strand which is similar to the conformation of LC/A observed in the context of holotoxin (Fig 1D) [23]. A major difference is that the β-strand interacts with the N-terminal region of the HC in the holotoxin while it is stabilized by pairing with the 250-loop in fLC/A. In contrast, the C-terminal peptide of the JPU-C10-bound fLC/A forms an amphipathic α-helix (F419–L429) in which the hydrophobic side is partially buried and facing the active site pocket while the hydrophilic residues are solvent-exposed (Fig 1C). We did not observe the electron density for residues R432 to K438 in the JPU-C10-bound fLC/A which are solvent-exposed and likely to have high flexibility. Therefore, the C-terminus of LC/A appears to adopt at least two distinct conformations, referred to as the β-form stabilized by JPU-B9 and the α-form captured by JPU-C10. In both conformations, residue C430 of fLC/A is solvent-exposed and thus likely available to form scrambled disulfide bonds with neighboring molecules in solution. This feature may explain why fLC/A is prone to aggregation and typically unsuitable for structural studies [24].
(A, B) The overall amino acid sequence identity (grey) and similarity (black) among BoNT/A or BoNT/B subtypes using A1 and B1 as the benchmarks. (C) The conservation of epitope sequences for selected LC/A- and LC/B-binding VHHs are shown. Green labels indicate comparable VHH binding to that specific subtype in comparison to A1 or B1, while red indicating weakened binding.
Botulinum neurotoxins are dangerous biothreat agents for which no post-intoxication antidote is available, and this results in a serious public health vulnerability. In a recent development, VHHs targeting LC/A or LC/B fused to an atoxic BoNT delivery vehicle were found capable of preventing death in mice, hamsters and primates that were treated post-symptomatically after receiving a lethal dose of BoNT/A or BoNT/B [9,10]. This opens a new avenue in the development of effective biomolecular antidotes for botulism in which sdAbs such as VHHs are delivered to the cytosol of intoxicated neurons where they inhibit the BoNT protease or target its degradation. In this study, we report the identification and comprehensive structural characterization of a large panel of VHHs as antidote candidates that bind to diverse epitopes on their LC/A or LC/B targets. Furthermore, studies with these VHHs revealed diverse mechanisms by which antibodies can inhibit BoNT protease activity. The findings should contribute significantly to the further development and refinement of biomolecular antidotes, as well as conventional drugs, for treating post-symptomatic botulism paralysis.
The availability of large panels of new VHHs binding to LC/A and LC/B identified in this study provide reagents able to probe a large portion of the surface of each protease for their functional role in SNARE protein recognition and cleavage (Figs 1 and 6). BoNT proteases are remarkably specific for their different SNARE protein substrates and this specificity is considered to be a result of the large region of interactions between these proteases and their substrates [15,33]. For LC/A, protease inhibiting VHHs were identified that bind to all four of the previously reported SNAP25-binding sites, called the α- and β-exosites and the anchoring sites I and II. Of note, JPU-A5 and JPU-C10 bind to the β-exosite of LC/A, which is a deep cleft at the active site of the protease and difficult to access by conventional Abs [37,38]. Our findings show that LC/A protease function is inhibited when a VHH can interfere with SNAP25 binding to any one of these four previously identified sites of interaction. Thus, this study strongly supports the essential roles of all four of the SNAP25-binding sites for LC/A cleavage to occur. In contrast to LC/A, we identified only two non-overlapping sites on LC/B at which protease inhibiting VHHs could bind and interfere with VAMP binding. This finding is consistent with reports that VAMP binds to LC/B through a smaller interface that is yet to be fully characterized [32,34].
Two alpacas were immunized with purified, full-length LC/A protein and two alpacas immunized with full-length LC/B as previously reported [8,12]. VHH-display phage libraries were prepared from immune PBLs obtained from each pair of immunized alpacas using standard lab procedures [42]. VHHs with affinity for native LC/A were selected by panning on LC/A immobilized by capture with the previously isolated ALc-B8 VHH [12] coated onto plastic, and this resulted in selection of the JPU series of VHHs (12] and this led to selection of several new LC/B-binding VHHs (JND series in 42].
VHHs identified in the discovery process were subcloned into the pET-32 expression vector, expressed as fusions to E. coli thioredoxin and purified by standard lab procedures [42]. Binding properties were assessed by dilution ELISAs employing LC/A or LC/B proteases (0.5 or 1 μg/ml) that were coated onto Nunc MaxiSorp or Costar tissue culture plastic, or captured to Nunc MaxiSorp plastic using various protease-binding VHHs (coated at 2.5 or 5 μg/ml). LC/A subtypes 1, 2, 3 and 4, or LC/B subtypes 1 and 8, were produced as strep-tag fusion proteins (as previously described in [7]) and employed in ELISAs in which the tagged proteases were captured to streptactin plates (IBA Lifesciences). Dilution ELISAs were performed as previously described [8] and apparent binding affinities under each of the ELISA conditions were assessed by estimating the EC50 values, i.e. the VHH concentration that resulted in 50% peak binding. Competition assays were done as first described in [6] with some modifications provided in [7].
14.Brunger AT, Breidenbach MA, Jin R, Fischer A, Santos JS, Montal M. Botulinum neurotoxin heavy chain belt as an intramolecular chaperone for the light chain. PLoS Pathog. 2007;3(9):1191–4. pmid:17907800; PubMed Central PMCID: PMC1994969.
41.Lam KH, Perry K, Shoemaker CB, Jin R. Two VHH antibodies neutralize botulinum neurotoxin E1 by blocking its membrane translocation in host cells. Toxins. 2020;12(10). pmid:32992561.

(A) Scattering profiles of BoNT/Ai at pH 8.0 (purple) and 6.0 (green). (i) Curve fitting of data at pH 8.0 with the crystal structure of free form BoNT/A. (ii) Curve fitting of data at pH 8.0 with the SAXS model determined by rigid body refinement. (iii) Superimposition of SAXS profiles at pH 8.0 and 6.0. Constant subtractions used in the rigid body refinements were taken into account. (iv) Curve fitting of data at pH 6.0 with the crystal structures of BoNT/A in the free (solid line) and the complex (dash line) forms. (v) Curve fitting of data at pH 6.0 with the SAXS model determined by rigid body refinement. All curve fittings were performed using the program CORAL [20]. (B) Kratky plots of the experimental profiles and theoretical curve of BoNT/Ai. (C) Pair distance distributions P(r) of BoNT/Ai at pH 8.0 and 6.0. Theoretical curves are shown as well. (D) The SAXS model of BoNT/Ai at pH 8.0 determined by rigid body refinement. The HC domain was refined as a rigid body (shown as brown tubes). Crystal structure of the free form BoNT/A is superimposed onto the LC–HN domain, with the HC domain labeled in purple tubes. The position of reconstructed loop is indicated by an arrow. (E) The SAXS model of BoNT/Ai at pH 6.0 with the HC domain labeled in cyan. (F) Comparison of the SAXS models between pH 8.0 and 6.0.
Songhai Tian, Xiaozhe Xiong, Ji Zeng, Siyu Wang, Benjamin Jean-Marie Tremblay, Peng Chen, Baohua Chen, Min Liu, Pengsheng Chen, Kuanwei Sheng, Daniel Zeve, Wanshu Qi, David T. Breault, César Rodríguez, Ralf Gerhard, Rongsheng Jin, Andrew C. Doxey, Min Dong

This work was partly supported by National Institute of Allergy and Infectious Diseases (NIAID) grants R01AI139087, R01AI158503, and R21AI156092 to RJ. NE-CAT at the Advanced Photon Source (APS) is supported by a grant from the National Institute of General Medical Sciences (P30 GM124165). Use of the APS, an Office of Science User Facility operated for the U.S. Department of Energy (DOE) Office of Science by Argonne National Laboratory, was supported by the U.S. DOE under Contract No. DE-AC02-06CH11357.
3. CDC. Antibiotic resistance threats in the united states, 2019. U.S. Department of Health and Human Services C (2019). Available at: https://www.cdc.gov/drugresistance/biggest-threats.html.
22. Chen B, Liu Z, Perry K, Jin R. Structure of the glucosyltransferase domain of tcda in complex with rhoa provides insights into substrate recognition. Sci Rep (2022) 12(1):9028. doi: 10.1038/s41598-022-12909-8
32. Kroh HK, Chandrasekaran R, Rosenthal K, Woods R, Jin X, Ohi MD, et al. Use of a neutralizing antibody helps identify structural features critical for binding of clostridium difficile toxin tcda to the host cell surface. J Biol Chem (2017) 292(35):14401–12. doi: 10.1074/jbc.M117.781112
49. Chen P, Jin R. Receptor binding mechanisms of clostridioides difficile toxin b and implications for therapeutics development. FEBS J (2021). doi: 10.1111/febs.16310
51. Kroh HK, Chandrasekaran R, Zhang Z, Rosenthal K, Woods R, Jin X, et al. A neutralizing antibody that blocks delivery of the enzymatic cargo of clostridium difficile toxin tcdb into host cells. J Biol Chem (2018) 293(3):941–52. doi: 10.1074/jbc.M117.813428
59. Lam K-H, Perry K, Shoemaker CB, Jin R. Two vhh antibodies neutralize botulinum neurotoxin E1 by blocking its membrane translocation in host cells. Toxins (Basel) (2020) 12(10):616. doi: 10.3390/toxins12100616



 8613371530291
8613371530291